
<aside>
🍁 基因过表达技术的缺陷:过表达未能反映出基因的真实功能;变异体并不适用于每个蛋白
</aside>
用显微注射针将线性化的外源 DNA 片段直接注入小鼠受精卵的原核中,使外源基因整合到小鼠基因组中,从而获得转基因小鼠。
因此随机转基因被称作是「简单粗暴,不清不楚」:转基因随机整合到基因组,敲入位点不清楚,拷贝数不清楚。
虽然我们可以利用 DNA 转座子系统将外源基因敲入在转座酶识别位点,进而提高转基因的表达阳性率,但依然避免不了随机转基因随机整合的缺点。
<aside> 🍁 表达比较活跃的基因区域通常不会被异染色质化(染色质处在固缩的状态),因此在这个区域定点插入外源DNA,在各组织中表达的可能性都非常高。然后从中筛选出对小鼠发育和繁殖无害的位点作为敲入位点,从而可以获得定点过表达的小鼠。因此定点转基因被称作是「一清二白」的过表达神器,
一清:整合位点清楚
二白:
1)整合拷贝数明白;
2)不需要建系,后代小鼠基因型明白。更重要的是:外源基因表达有保证!所以说定点转基因的关键就在于“定点”,而定点整合位点--SafeHarbor的选择尤为重要。
可以安全地进行外源基因插入并高效表达的点位:
Rosa26、AAVS1、CCR5、HPRT、H11、Col1a1、TIGRE

20 世纪 90 年代末 Philipe Soriano 及其同事发现,一个被命名为 ROSAβgeo26 的随机转基因小鼠品系在所有组织中都能检测到高水平的β半乳糖苷酶表达。他们对这种转基因小鼠进行了基因插入的定位,确定了在第6号染色体的基因整合,也就是我们现在常说的Rosa26位点。
Fig.1 ROSA26基因组区域图
在基因定点条件性过表达小鼠模型的构建中,利用Cre-LoxP系统对目的基因进行可诱导表达调控。构建Rosa26-(SA/pCAG)-loxp-Stop-loxp-cDNA-pA 重组载体,将条件性过表达结构插入到Rosa26基因intron1中。该类型Knockin小鼠可与各类表达Cre重组酶的工具鼠杂交,获得组织特异性表达外源基因的条件性过表达小鼠模型。
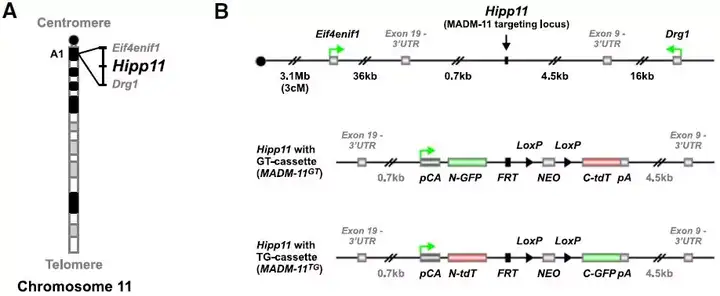
Fig.3 H11基因组区域图

H11位点(也叫Hipp11)位于小鼠第11号染色体,是 Eif4enif1与Drg1 这两个基因之间的一个位点,由 Simon Hippenmeyer于2010 年发现并命名。由于H11位点位于两个基因之间,故外源基因插入后影响內源基因表达的风险很小。同时H11位点所在的Eif4enif1与Drg1这两个基因的侧翼序列区域具有广泛的空间和时间EST(expression sequence tag)表达模式,能够使整合在此的外源基因受指定启动子的驱动而稳定表达。该位点的纯合敲入小鼠可正常发育和繁殖。因此成为可与Rosa26位点相媲美的新「Safe Harbor」。
和Rosa26位点的过表达模型一样,在H11位点我们也可以采用CRISPR/Cas9技术构建条件性基因过表达小鼠模型,即广泛型强启动子pCAG与外源目的基因cDNA之间用loxP-STOP-loxP结构隔开,只有与组织特异性Cre工具鼠交配后,才会在特异细胞或组织中表达目的基因。
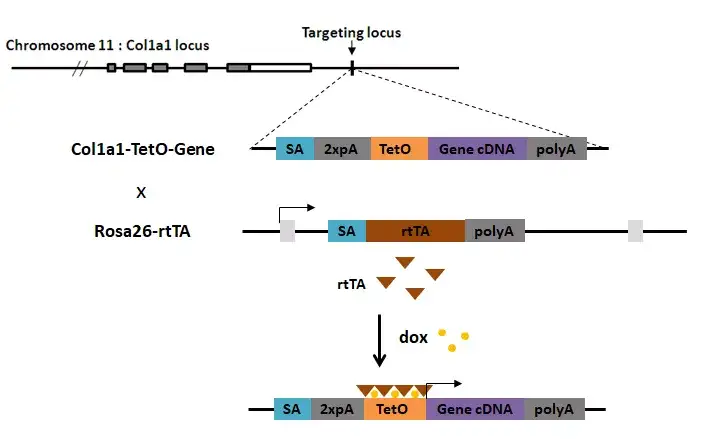
Col1a1位点是2006年由Caroline Beard首先使用作为四环素诱导调控外源基因表达的整合位点(四环素诱导表达 Tet-on和Tet-off)。Col1a1位点实际上位于第11号染色体的Col1a1基因3‘UTR下游约500bp左右。在四环素调控模型中,在TetO启动子(只有结合tTA才会启动后续基因的表达)之前往往加上SA-polyA来阻隔上游启动子的影响,防止不依赖于四环素的转录调控。
Fig.5 Col1a1位点四环素调控可逆过表达构建策略
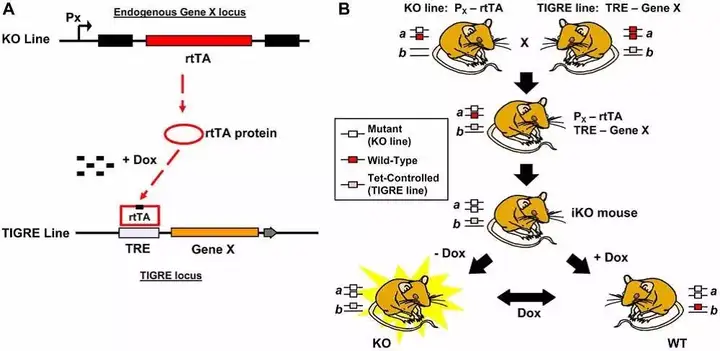
Fig.6 TIGRE line诱导敲除小鼠构建策略

TIGRE是 tightly regulated 的缩写,也是一个适用于四环素调控的整合位点,位于小鼠第9号染色体AB124611 和 Carm1两个基因之间。和Col1a1位点类似,最好在TetO-cDNA-polyA两侧加上**隔离子(染色体上能够保护基因免受其外部增强子和非活性染色质结构影响的DNA 序列)**来减少不依赖于四环素的本底表达。
The ApoE TIGRE locus contains an exogenous copy of ApoE cDNA driven by TRE and flanked by four copies of chicken b-globin insulator (ins) sequences, two on each side, and a PGK promoter (P) driven loxneo selection marker.